1995 年的诺贝尔奖,颁给了一只果蝇背后的科学,1995 年,诺贝尔生理学或医学奖授予三位科学家Edward B. Lewis、Christiane Nüsslein-Volhard 和 Eric Wieschaus。他们的研究对象不是疾病,也不是药物,而是一个更基础、也更困难的问题:胚胎早期,身体结构是如何被“设计”出来的?答案,就藏在果蝇胚胎里。
为什么偏偏是果蝇?
黑腹果蝇(Drosophila melanogaster)是生命科学中最经典的实验动物之一,原因很现实:发育快:10 天完成一代;后代多:一次实验可获得数百枚胚胎;遗传学成熟:基因操作工具完善;胚胎透明、结构清晰:非常适合观察早期发育;更重要的是——果蝇的发育调控逻辑,与人类高度保守。研究果蝇,其实是在“低成本地理解我们自己”。
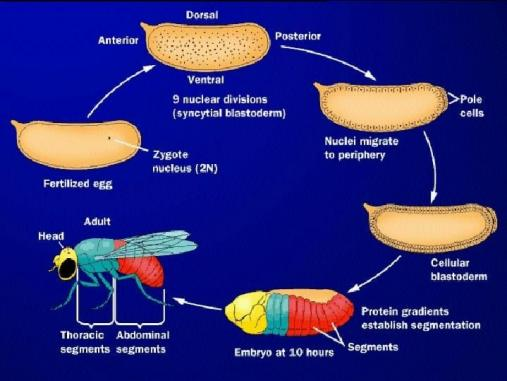
果蝇胚胎:一个“反常识”的开始
果蝇胚胎的早期发育,非常不像我们想象中的“细胞分裂”。在最初几个小时里,它会经历多轮核分裂,却不分裂成细胞,形成一个“多核共用空间”的合胞体胚胎。这意味着什么?调控发育的蛋白质,可以在整个胚胎中自由扩散;形成浓度梯度,告诉细胞“你在哪个位置”;这是后续身体轴线和分节形成的关键前提。
一场教科书级的实验:大规模突变筛选
真正改变教科书的,是一项近乎“疯狂”的实验设计。科学家使用化学诱变剂,随机制造果蝇基因突变,然后做一件事:看胚胎长坏了哪里。
他们建立了约 5800 条果蝇稳定遗传品系,其中 2600 条为胚胎致死突变品系,研究人员逐一检查这些胚胎的角质层结构。
这不是“挑几个基因研究”,而是一次地毯式搜索发育必需基因的工程。
结果有多震撼?
最终,他们发现:约 600 个突变体表现出清晰的发育异常;这些突变集中对应约 120 个关键基因;而且这些基因并非各自为政,而是形成了一条完整链条:母源基因:建立胚胎坐标;缺口基因:划定大区域 ;对规则基因:确定分节节律节段;极性基因:精细修饰边界。这是一套高度程序化的“身体建模流程”。
最经典的画面:长错位置的器官
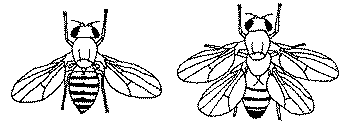
其中最具视觉冲击力的,是 Lewis 研究的同源异型基因。这些基因不决定“长不长器官”,而是决定器官长在什么位置。当某些基因发生突变时,果蝇会在本该长平衡棒的位置,长出一对完整的翅膀。这告诉我们一个颠覆直觉的事实:身体结构不是一步步“拼出来的”,而是被基因提前“规划好”的。
实验动物,也有“标准写法”
在实际研究中,科学家会明确记录实验动物信息,例如: 物种:Drosophila melanogaster;常用品系:Canton-S、w1118;胚胎收集规模:常以约 500 只成虫组成产卵群体;实验对象:同步发育的早期胚胎;这些细节,是实验可重复性的基础,也是现代科研的底线。
一只果蝇,改变了我们对生命的理解
这项研究的意义,远不止果蝇本身。它第一次让人类清楚地看到:从基因到身体结构,并不是混乱堆砌,而是精密执行的程序。今天我们理解的先天缺陷、器官错位、甚至部分肿瘤机制,都能在这套发育逻辑中找到源头。而这一切,始于一只不起眼的小果蝇。
写在最后
实验动物的价值,不在于“像不像人”,而在于 它是否能让我们更早、更清楚地看见生命的底层规律。
参考文献:
Nüsslein-Volhard, C., & Wieschaus, E. (1980). Mutations affecting segment number and polarity in Drosophila. Nature, 287(5785), 795-801.
Lewis, E. B. (1978). A gene complex controlling segmentation in Drosophila. Nature, 276(5688), 565-570.
Driever, W., & Nüsslein-Volhard, C. (1988). The bicoid protein is a positional cue in the early Drosophila embryo. Cell, 54(1), 95-104.

