今天想和大家分享的是一篇关于癌症起源的文章(Cell Rep,9.4233)。癌症很可怕,但比癌症更可怕的是癌症转移,癌细胞拥有无限的生长能力,增殖能力和转移能力。一旦发生转移,癌细胞就会侵入人体其他组织器官,导致器官衰竭甚至死亡。此外,在癌症转移发生后,很多癌症治疗方法都会受到限制,治疗难度大大提高。然而,现阶段研究对 “转移细胞是如何产生的” 这一问题尚不清楚。引起转移的细胞必须首先在原发肿瘤内获得促转移状态,再参与迁移机制,并最终在其他地方形成继发肿瘤块。那么这种促转移状态以及转移细胞又是如何产生的呢?就让我们一起带着这个问题开始今天的文献学习之旅吧。
日内瓦大学的科学家们通过寻找诱导最近表征的促转移状态,作为转移起源的替代物,并进一步探究细胞死亡诱导疗法是否会诱导人类结肠癌细胞的促转移状态。最终发现一类获得促转移状态的濒死细胞(PAME),并发现其拥有在体内形成远端转移的能力。这些 PAME 细胞表现出细胞因子风暴,内质网 (ER) 应激和核重编程能力增强等特点。PAMEs 诱导邻近肿瘤细胞成为 PAME 诱导的迁移细胞(PIMs): 其同样拥有高度迁移能力,重现细胞因子风暴并增强 PAME 迁移(图解摘要)。因此,研究者认为濒死细胞可以通过内质网应激调节、转移重编程和细胞因子风暴获得促转移能力。
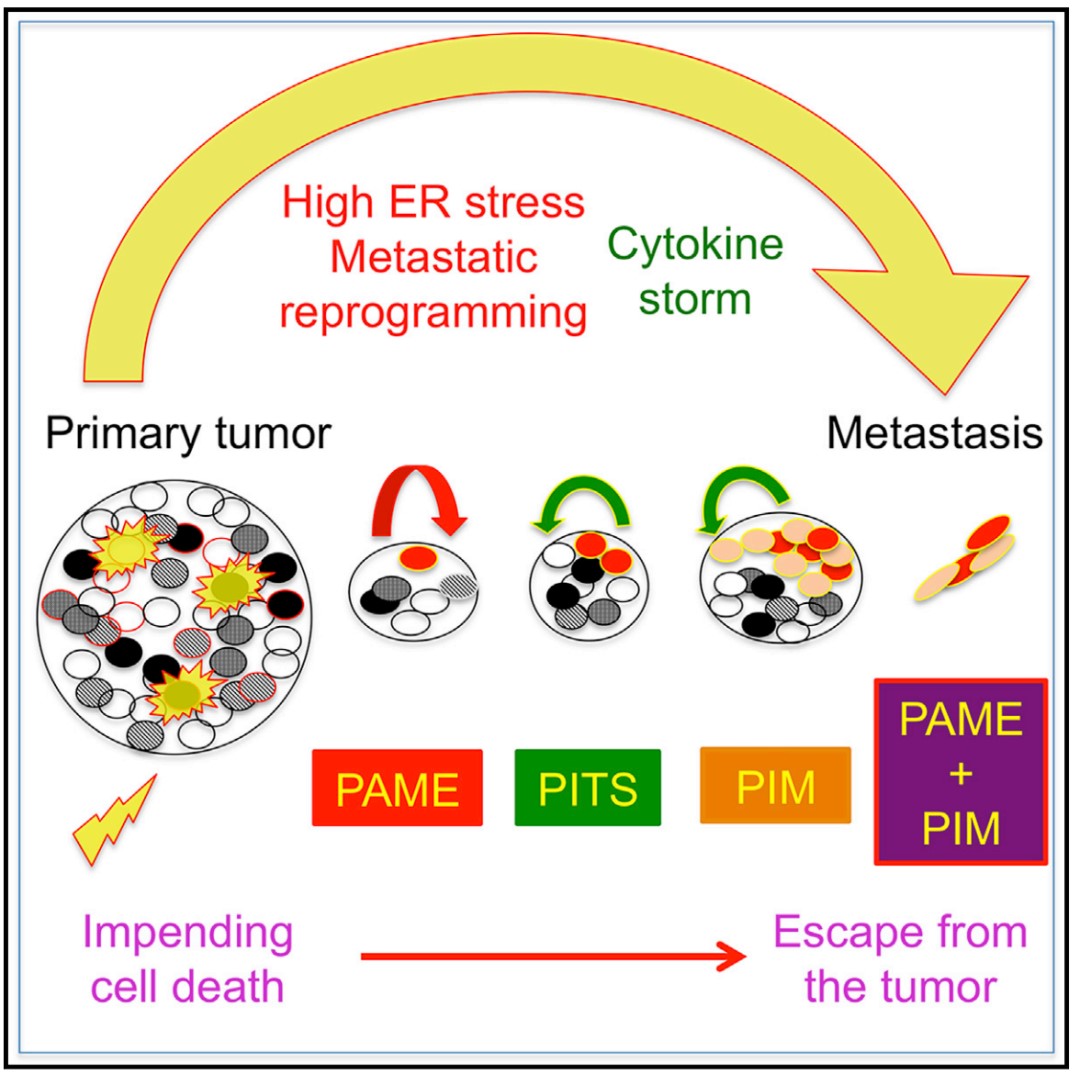
图解摘要
On the origin of metastases: Induction of pro-metastatic states after impending cell death via ER stress, reprogramming, and a cytokine storm
转移的起源:通过内质网应激、重编程和细胞因子风暴诱导细胞濒临死亡后的促转移状态
1.濒死细胞活下来,就成了我:PAME
对异质结肠癌 CC14 细胞进行细胞致死 STS 处理(激酶抑制剂十字孢碱触发晚期凋亡细胞的存活),6 h 后用凋亡抑制剂 Q-VD-OPh + DIDS 进行阻断(图 1A-B)。仅用 DMSO 或 BLOCK 处理的细胞 (对照组) 不受影响 (图 1B),而用 STS 处理的细胞 100% 死亡并出现凋亡表型(图 1B-C)。STS + BLOCK 处理 5 天后部分 PAMEs 被挽救并出现增殖(图 1C)。PAMEs 经慢病毒转染后均为 lacZ+,并经 XGal 染色鉴定(图 1D)。而单独的 BLOCK 无效,PAMEs 的迁移细胞数量增加 12 倍(图 1D)。在其他结肠癌 CC36,Ls174T, DLD1 等细胞株中均可观察到相似现象(图 1E-F)。将 CC14 PAME lacZ 移植到裸鼠中(图 1H),与 DMSO/DMSO 或对照(DMSO/BLOCK) 组相比,肺远端转移瘤分别增加 3.4 倍和 1.5 倍(图 1H),表明存在内源性凋亡细胞的挽救现象。从 CC14 PAME 原发肿瘤中移植的细胞显示出 2 倍的迁移能力(图 1I),表明 PAME 后代在多次分裂后仍保持着较强的转移能力。
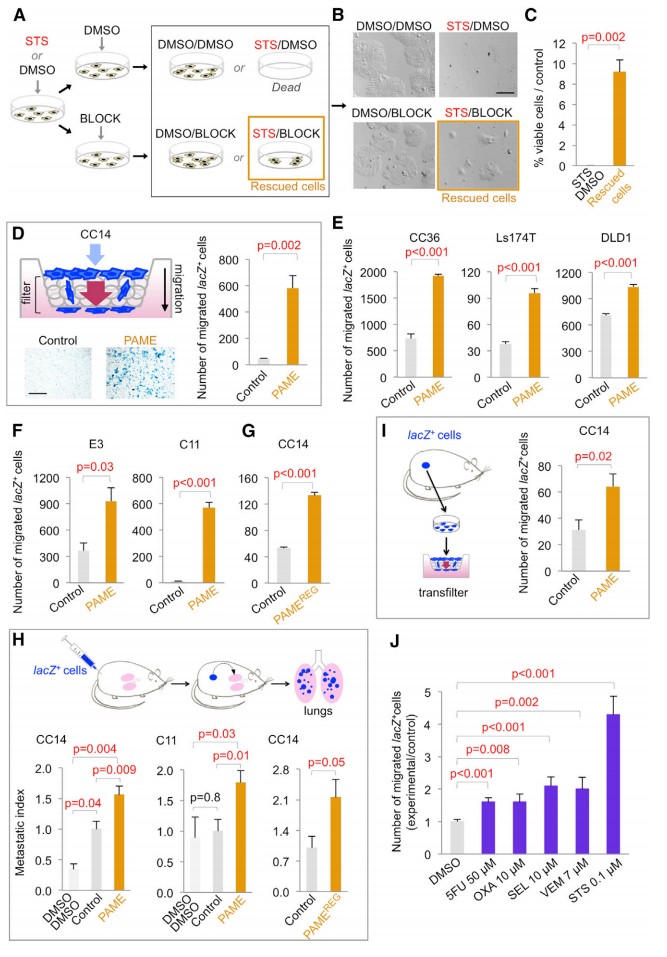
图 1. PAMEs 表现出稳定、增强的迁移和转移能力
2.凋亡阻断剂相关的多种药物诱导 PAME
通过 regorafenib 和 BLOCK 处理 4 天后,细胞数量达到原来的 30% - 40%。经过 2 天的恢复和扩展后,PAMEs REG 显示出 2.5 倍的迁移能力 (图 1G)。此外,用非 apocalyptic STS 处理,然后在不含 BLOCK 的环境培养 7 天后,细胞的迁移能力提高 4 倍,其他临床相关化疗药物处理后,细胞迁移能力也表现出不同程度的增强 (图 1J)。
3. PAMEs中分泌性、干性和转移性重编程因子表达增强
研究者进一步对 PAME-CC14(STS PAMEs)和对照细胞进行 RNA-seq 测序分析,在 STS PAMEs 中细胞因子信号转导、炎症 / 免疫反应和细胞外基质组织相关基因表达上调,有利于细胞因子风暴的形成。额外的 qRT-PCR 分析显示 SNAIL1、ZEB1 和 VIM 在 PAMEs 中升高 2 倍或更多 (图 2B),提示在此过程中 EMT 能力增强。此外,所有被检测的结肠癌 PAMEs 都具有高水平表达的干细胞标记物 NANOG/NANOGP8(图 2B),并且大多数也表达高水平的转移性重编程因子 OCT4、SOX2 和 KLF4 (图 2B)。
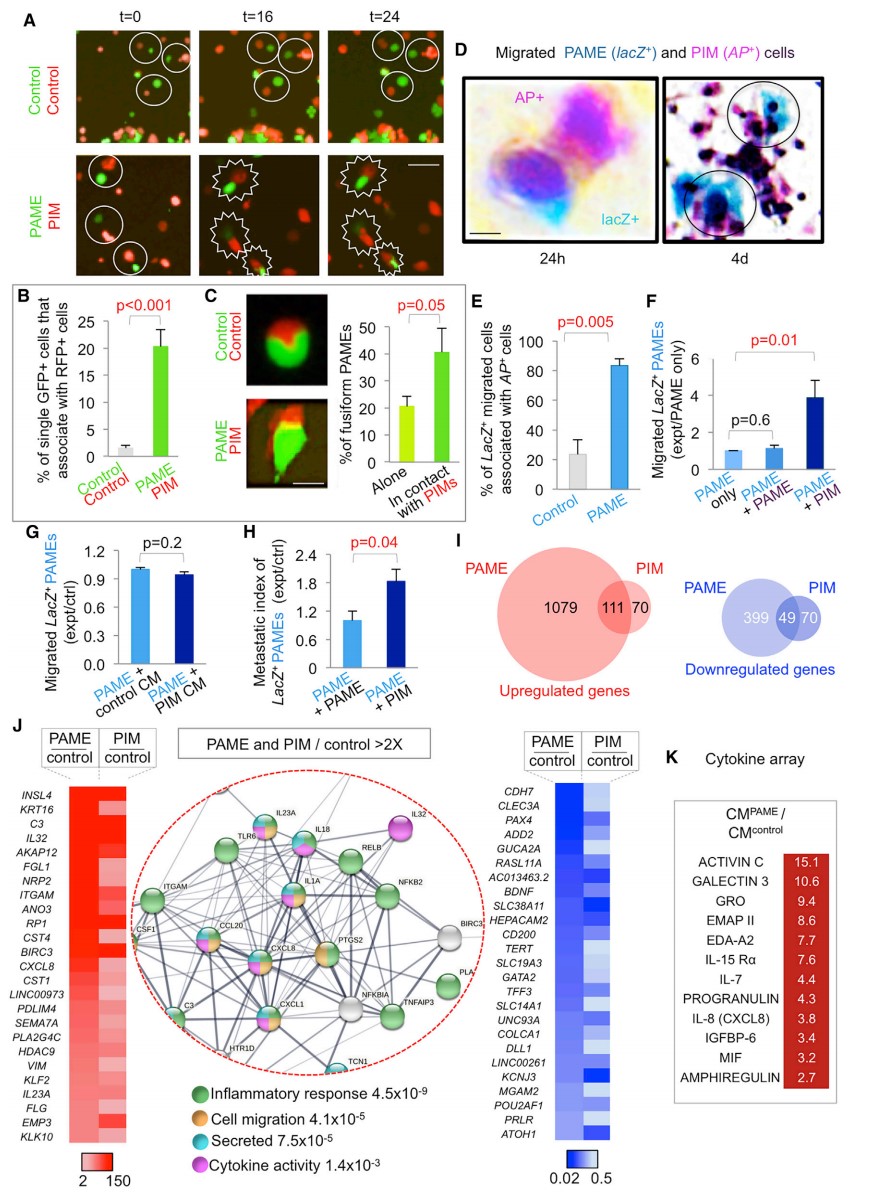
图 2. PAME 转录组分析和 PIM 诱导
4. PAMEs诱导邻近癌细胞成为高度迁移的 PIMs
分别将 RFP+CC14 细胞与对照细胞和 PAMEs 共培养,相比对照组,与 PAMEs 共培养的细胞其迁移能力增加 9 倍 (图 2C-D)。被 PAMEs 诱导获得高度迁移能力的肿瘤细胞被称为 PIMs (PAMEs 诱导迁移细胞)。PAMEs GFP 和 PIMs RFP 的基因表达分析表明 PAMEs 中上调的重编程 / 干性特征在 PIMs 中缺失(图 2E)。然而,两种细胞在显著上调和显著下调的 top 基因中表现出明显重叠(图 2E)。为测试 PIM 诱导扩散的可能性,研究者使用 C11 细胞进一步检测 PIM (PIMs1) 诱导更多 PIM (PIMs2)的能力。PAMEs 诱导的 PIMs1 被 FACS 分离,其迁移能力增加 26 倍(图 3A-B)。此外,经 PIMs1 诱导后生成的 PIMs2 同样保持高度迁移能力,虽然这种能力稍低于 PIMs1(图 3C-D)。此外,研究者发现在迁移能力越强的细胞中,INSL4 和 IL32 表达越高(图 3E-F)。PIM 诱导在传播过程中迁移能力逐渐下降。
5. PITS分泌诱导 PIM是一种常见现象
研究者预测 PIM 诱导会分泌细胞因子风暴 (PITS),因此进一步测试 PAME 培养基对 PIM 的诱导(图 3G)。在研究者发现来自不同细胞甚至多个细胞的 PAMEs 培养基(CM) 诱导的 PIMs 相比对照 CM 处理的同类型细胞均发生不同程度的增加 (图 3H-J)。
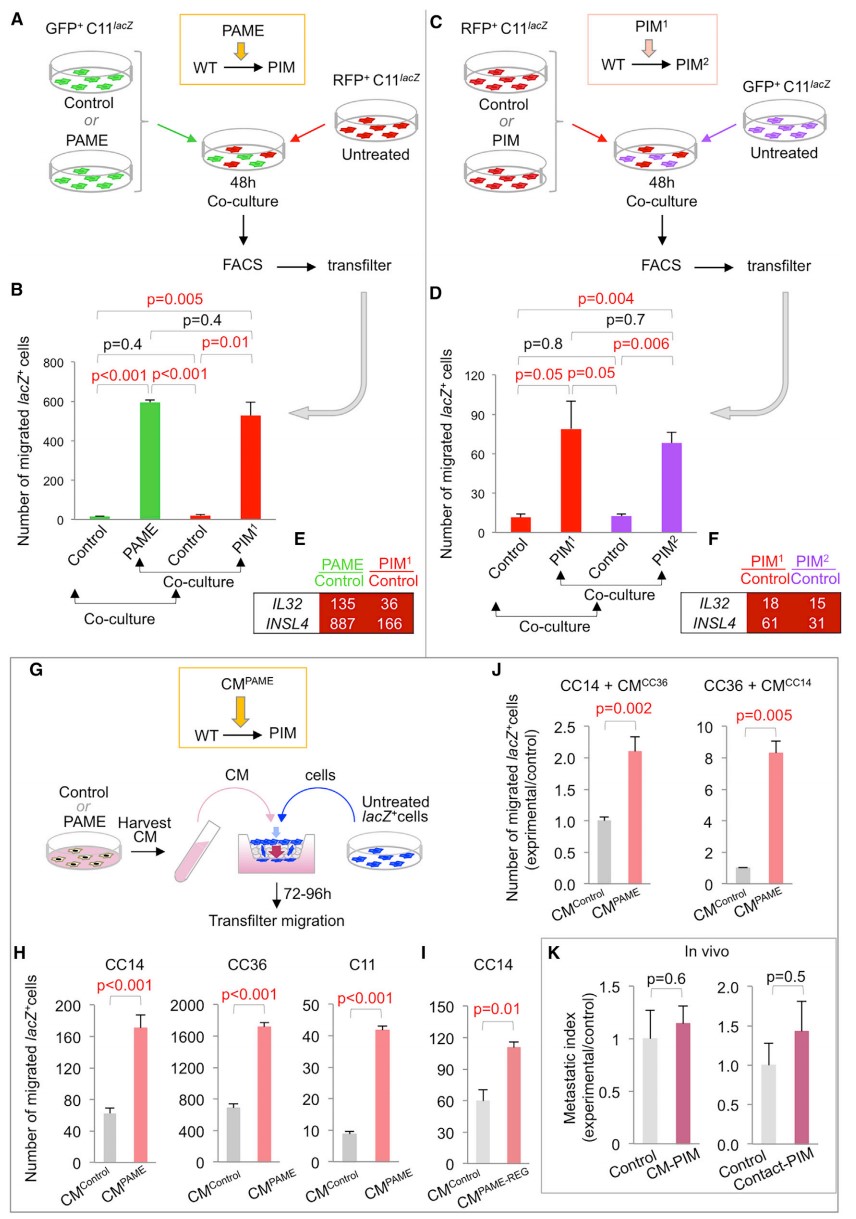
图 3. PIM 诱导和条件培养基
6. PIMs与 PAMEs关联并发生共同迁移
与对照组相比,将 CC14 PIMs 移植到裸鼠中并没有增加其转移能力 (图 3K)。因此研究者推测 PIMs 可能会帮助 PAMEs 转移。标记共培养物的延时照片分析显示 PAME-PIM 细胞的关联频率是对照 - 对照细胞的 10 倍 (图 4A 和 4B)。这种关联很密切,在共焦显微镜中出现黄色重叠区 (图 4C)。梭形细胞形态(通常与主动迁移有关)在 20% 的 PAMEs 中存在,但在对照细胞中非常少见 (图 4C)。与 PIMs-RFP 相关的 PAMEs-GFP 表现出这种表型的频率是单独 PAMEs 的两倍 (图 4C)。对 24 小时后穿过过滤器的细胞进行染色,当超过 95% 的细胞为单一细胞时,显示 CC14 PAME-lacZ 与 PIM-AP 混合后细胞比对照混合细胞增加 2.5 倍 (图 4D),并且这种趋势可以保持 4 天。以上结果表明 PAMEs-lacZ 与 PIM-AP 的相关性比对照组 lacZ、AP 混合物增强 4 倍 (图 4D-E)。此外,与 PAMEs 单独培养或 PAMEs- lacz 与细胞分选后的 PAMEs- GFP /AP 共培养相比,PAMEs-PIMs 共培养细胞的的迁移增加 4 倍 (图 4F)。PIM 培养基对 PAME 迁移没有影响 (图 4G)。当 PAME 与 PIM 共同存在时的转移效率比 PAME 与 PAME 共同培养时提高 1.8 倍 (图 4H)。

图 4. PAMEs/ PIM 的关联和迁移及常见细胞因子风暴的识别
7. PIM和 PAME转录组的相似性
与对照组相比,PAMEs 和 PIM 存在显著交叠的差异表达基因,其中炎症反应、细胞因子活性、细胞迁移的正向调节以及分泌因子相关基因在 PAMEs 和 PIM 中同时上调,并且存在频繁的蛋白质互作 (图 4I-J)。此外,与对照组相比,12 个细胞因子蛋白在 CC14-PAME 培养基中表达丰度增加 2 倍(图 4K),表明 PITS 含有多种细胞因子和分泌因子。
8. PITS与 CXCL8、MIF、EGFR、COX2、IL32和 INSL4的活性相关
CXCL8 在 PAMEs 和 PIM 中上调,在蛋白质互作网络和细胞转移过程中发挥关键作用。研究者在 CC14 细胞中加入 CXCL8 配体后,其迁移细胞数量增加 (图 5A)。相比之下,CXCL1 配体加入后迁移细胞数量下降 (图 5A)。为抑制内源性 CXCL8 功能,研究者将一种 CXCR1/CXCR2 受体阻断剂添加到培养皿中,预处理 1 小时后发现 PAMEs 诱导的 PIM 迁移量减少 2 倍 (图 5B)。
AZD-5069 是另一种 CXCR2 的拮抗剂,但 AZD-5069 处理后未观察到迁移变化 (图 5B),表明 CXCL8 通过 CXCR1 发挥作用。由于 0.1-1 ug/mL 的 CXCL8(图 5A) 不能表现出 PITS 的强度(图 3G-H),研究者进一步在 PAMEs 中测试其他上调因子的影响。ISO-1 处理 PAME 诱导 PIM 后,PIM 迁移率降低 30%(图 5B)。在 PAME 中 AREG 上调后迁移能力增加(图 5A),而用西妥昔单抗阻断其受体 EGFR 的活性会导致 PIM 迁移的减少(图 5B)。
IL32 和 INSL 在 PAMEs 和 PIM 中上调,参与迁移和侵袭调节过程。IL32 诱导迁移能力增强 (图 5A)。在 CC14 细胞中,通过 cDNA 转导直接表达 INSL4 可使细胞的迁移行为增强 2 倍 (图 5A),这与 CXCL8 无关。与 CXCL8 不同,在药理学上无法阻断 INSL4 作用 (图 5B),而且由于其在 PAMEs 中的诱导作用非常高,因此难以充分抑制 INSL4,这促使研究者构建 CRISPR-Cas9 双突变克隆,删除其启动子 ATGs。在体外,INSL4 和 IL32 突变体使迁移能力和远端转移能力显著下降 (图 5C)。
9. PAMEs和 PIM显示单细胞定义的促转移状态
研究者先前的 scRNA-seq 工作分析识别出一类 METSP(促转移细胞亚群,图 5D),CXCL8、INSL4 和 IL32 在 METSP 中高表达 (图 4J)。为进一步确定 PAMEs 和 PIM 是否表现出 METSP 促转移状态,研究者将在 RNA-seq(也在 scRNA-seq 中检测到) 中常见的 PAME/PIM 上调和下调基因映射到四个先前定义的 scRNA-seq 簇 (图 5D- E)。PAME/PIM 上调基因主要在 METSP 中表达,而下调基因则映射到非 METSP 中 (图 5E)。在 PAME/PIM 中上调且显著富集到 METSP 中的基因主要与浸润,细胞因子活性等生物学过程相关 (图 5F)。
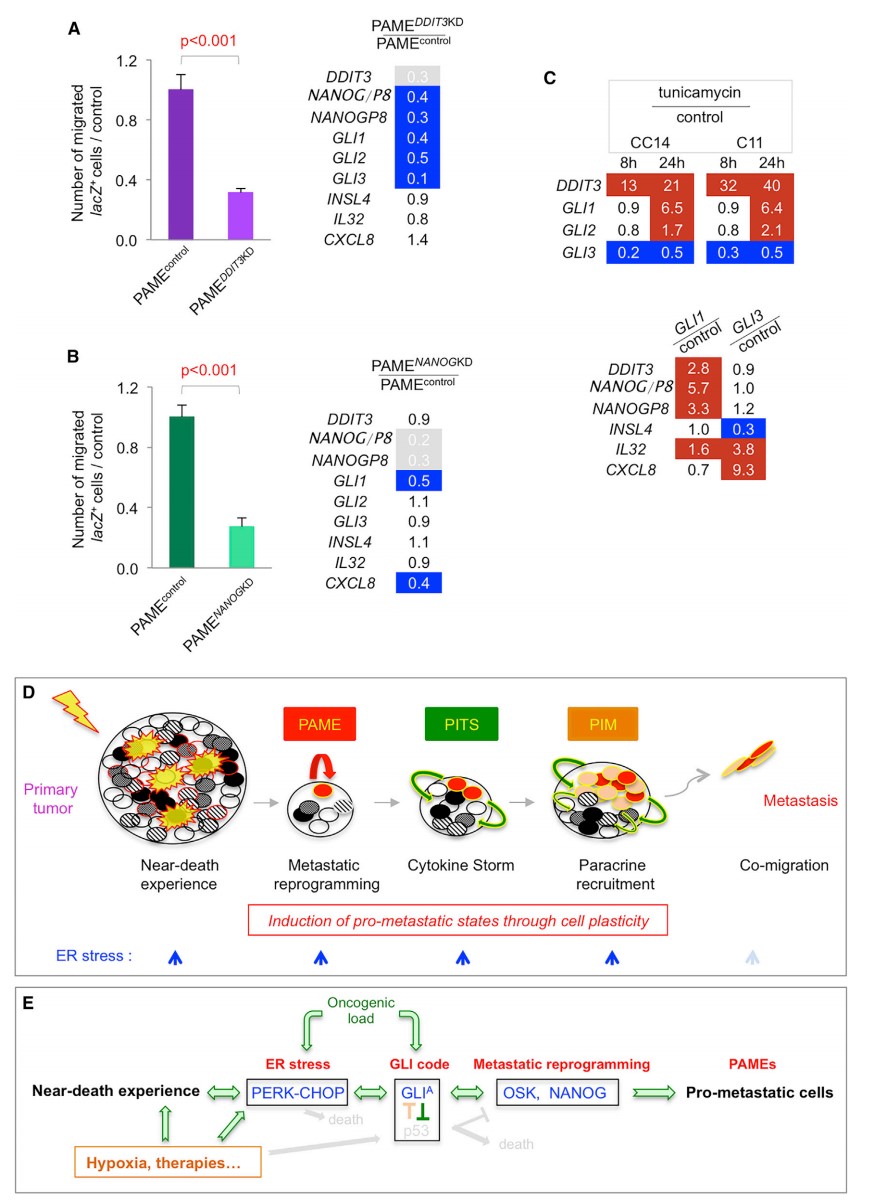
图 5.PAMEs 和 PIMs 与多功能细胞因子风暴相关,并表现出促转移
1. PAMEs 与内质网应激
METSP 细胞内质网应激相关基因,如 DDIT3(编码 CHOP)、HSPA5(编码 BIP)、ATF 等表达增强 (图 6A)。一致地,研究者发现在 PAME-CC14 和 PAME-C11 中,发生剪接的 XBP1、DDIT3 和 HSPA5 的表达水平有所提高(图 6B)。鉴于内质网应激和转移之间的联系,研究者进一步测试高度内质网应激对驱动迁移能力的影响。经内质网应激诱导处理后的 CC14 和 E3 细胞中 DDIT3 和 HSPA5 表达升高,迁移能力降低(图 6C,蓝条)。研究者进一步基于 PERK 抑制剂(PERKi;GSK2606414) 测试内质网应激相关通路 PERK-ATF4- CHOP 对迁移的影响。PERKi 处理增强其迁移能力,降低 DDIT3 和 HSPA5 表达(图 6C 和 6D), PERKi 和内质网应激诱导处理相互平衡(图 6C)。此外,迁移活跃的 PAMEs-C11 和 PIM 中内质网应激标记物水平降低(图 6E),这与迁移的 CC14 细胞中 HSPA5 下调一致,但其他 METSP 基因保持不变(图 6E)。PERKi 阻断 PREK,进一步增强 PAME-CC14 和 PAME-C11 的细胞迁移(图 6F-G)。

图 6. 内质网应激调节 PAME 迁移能力
1. CHOP、NANOG 和 GLI 参与 PAMEs
CHOP 是 PERK 的下游因子,研究者通过敲除 DDIT3 验证其对迁移的影响。DDIT3 敲除后 PAME 细胞的迁移能力显著下降 (图 7A)。然而,除 NANOG/P8 和 GLI1 没有明显表达增加外,DDIT3 敲除 PAME 中的 PAME 特征仍然存在 (图 7A)。因此,研究者推测 CHOP 对于转移性 PAMEs 重编程是必要的。研究者进一步直接测试 NANOG 的作用,发现 NANOG 在癌细胞中是由转移性重编程诱导以及增强转移状态所必需的,并与 GLI1 发挥协同作用。NANOG/NANOGP8 敲除未观察到 GLI1 和 CXCL8 的上调以及转移能力的增强 (图 7B)。
上述结果提示 GLI 基因可能在内质网应激和转移性重编程 / NANOG 之间发挥关联作用。因此,研究者进一步探究内质网应激增强是否可以激活 GLI1。内质网诱导处理后 GLI1 表达升高 (图 7C)。GLI1 的表达导致 NANOG/P8 上调,IL32 增强 (图 7C),GLI3 影响 IL32 和 CXCL8,抑制 INSL4(图 7C)。因此,研究者认为主要是由 GLI1,而不是 GLI3 负责调节 PAME 特征和内质网应激水平的反馈,以及 DDIT3 表达升高 (图 7C)。
以上结果表明在被攻击后侥幸存活的人类结肠癌细胞,可以通过细胞重塑,诱导促转移状态。濒死细胞可以通过内质网应激调节、转移重编程和细胞因子风暴获得促转移能力。(图 7D-E)。
图 7. 通过 CHOP、NANOG 和 GLI 参与的 PAMEs 诱导转移发生
据报道,除非肿瘤过大,压迫到重要脏器以外,绝大部分癌症病人并不是直接死于原发肿瘤。然而,癌细胞转移会使脑,骨等重要器官收到侵害,数据显示 90% 的患者死于癌症转移。所以癌细胞的转移,对控制和治疗肿瘤是至关重要的。在本篇文章中,研究者将诱导最近表征的促转移状态作为转移起源的替代物。通过诱导濒死细胞并测试其转移、迁移能力,发现这些本应死亡的细胞会自我重新编程,然后呈现出更高的转移风险。此外,研究者还发现细胞濒死状态产生内质网应激,从而使 PAME 细胞得以发育以及细胞因子风暴产生。该研究为新的癌症转移治疗策略铺平道路,有利于防止某些由于治疗产生的促转移区域的发展。PAME 细胞可能是未来潜在的治疗和转移预防靶点。
以上研究主要是基于实验方法探索转移起源,那么在我们生物信息学分析中,又该如何定义影响癌症患者转移的关键因素呢?下面小编还想和大家分享一个探究人类癌症转移的数据库:HCMDB。
肿瘤转移是癌症患者死亡的主要原因。在过去十年中,高通量技术已经提供与癌症转移相关转录组变化的全基因组概括。许多微阵列和 RNA 测序研究致力于寻找不同类型癌症的转移相关表达模式。然而,这些表达数据仅仅被分散存储在各个数据库中,缺乏一个集成的数据挖掘平台,并对这些数据进行高效地分析挖掘。为便于对转移相关转录组数据的深入分析,研究者对这些转移相关的表达数据进行全面整合,并构建出 HCMDB(人类癌症转移数据库,http://hcmdb.i-sanger.com/index),在这里我们可以免费查询跨平台转移的转录组数据。
图 8. HCMDB 数据库
HCMDB 的大规模转录组数据主要来源于 GEO、SRA、TCGA 数据库。在当前版本的 HCMDB 中,包含来自超过 455 个实验的 29 种癌症类型。为注释这些潜在的转移相关基因,研究者从 7000 多篇发表的文献中收集 2183 个基因 (1901 个蛋白编码基因,24 个 lncRNA 和 203 个 miRNAs),构建易于使用的癌转移相关基因查询和浏览界面。
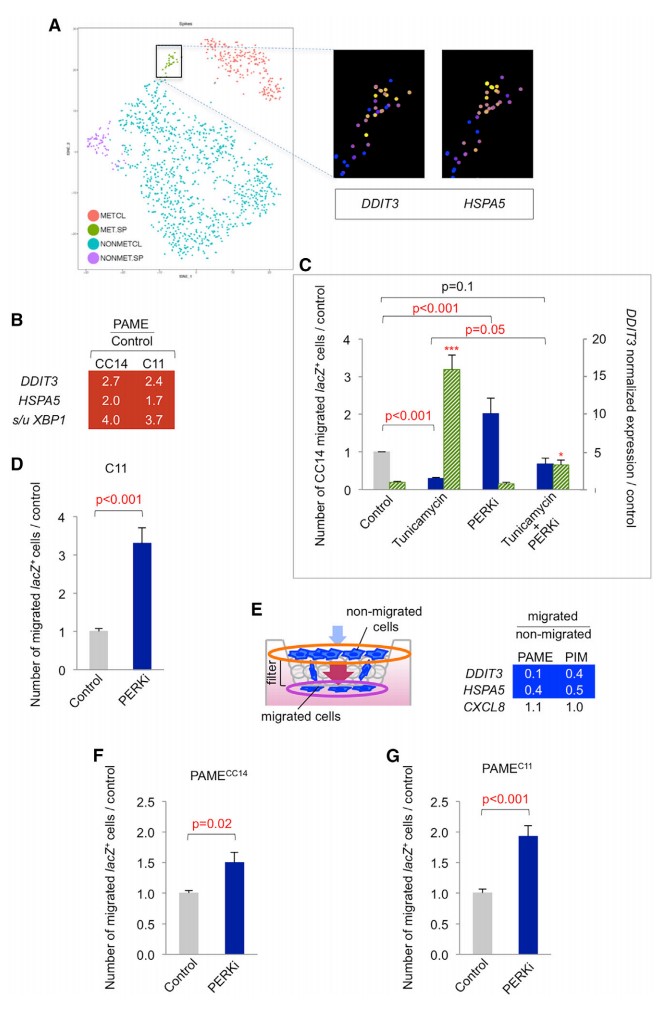
表 1. HCMDB 的数据资源
该数据库提供搜索功能,可以提供相关基因基本信息,以及在转移样本中的表达变化:
图 9. 检索功能
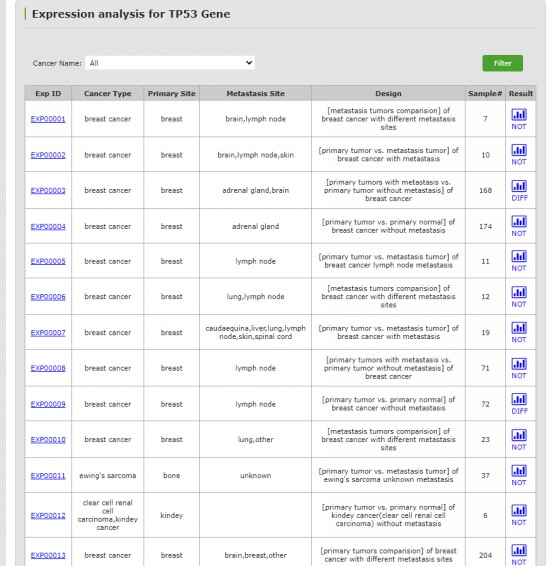
图 10. 转移相关基因的表达分析
此外,研究者还可以对不同数据中在转移与非转移样本中差异表达基因等进行检索,并提供可视化结果。

图 11. 差异表达基因及其可视化结果
当然,本文提供数据集合以及实验支持基因转移相关数据集下载。

图 12. 数据下载
除转录组差异外,近期发表于 CELL 的一篇文章也对癌症转移的基因组特征进行探究,该工作主要基于整合大规模基因组和临床数据,确定不同肿瘤类型转移模式和基因组改变之间的关联,为研究转移扩散的生物学基础提供宝贵的资源,并强调染色体不稳定性在癌症进展中的复杂作用,在这里我们就不重点介绍了,感兴趣的小伙伴们一定要自己去看哦。
图 13. 癌症转移的基因组特征
癌细胞固然极其 “可恨”,但是它顽强的意志品质却值得我们每个人学习。希望我们都能像“癌细胞” 一样顽强,哪怕是只有一线生机,也绝不放弃任何前进的希望。今天的分享到这里就结束啦,我们下次再见~
参考文献:
1.On the origin of metastases: Induction of pro-metastatic states after impending cell death via ER stress, reprogramming, and a cytokine storm
1. HCMDB: the human cancer metastasis database
2. Genomic characterization of metastatic patterns from prospective clinical sequencing of 25,000 patients
转载自公众号《生信人》,如有侵权,请联系删除。

