近几十年来,表皮激肽酶相关肽酶 (KLKs) 的功能鉴定引起了人们的极大兴趣。KLKs 由 15 种丝氨酸蛋白酶组成,它们的活性受复杂而精细的机制调控,包括蛋白水解激活级联、内源性抑制剂和环境因子。KLKs 参与各种活动,如皮肤炎症、伤口愈合、瘙痒、抗菌活性和病毒易感性。KLKs(KLK5 和 KLK7)的主要作用之一是生理脱屑。这两种蛋白酶也参与伴有屏障异常的炎症性皮肤病的发展。其他的 KLKs(包括 KLK6 和 KLK8)也在健康和患病皮肤 (如牛皮癣) 中的功能和意义是一个令人兴奋但相对未开发的领域。这篇文章中我们阐明了KLKs 与肿瘤微环境的发生发展之间的关系并且将有助于开发疾病特异性生物标志物和新的治疗策略。

激肽释放酶相关肽酶(KLKs)
KLKs 是一种蛋白水解酶,人类激肽酶相关肽酶 (KLK) 家族由 15 个高度保守的丝氨酸蛋白酶组成,它们由人类基因组中最大的不间断蛋白酶基因簇编码。可以调节生物活性分子的多种功能,包括激素和生长因子、膜受体和参与癌症进展和转移的细胞外基质结构。由这些肽酶产生的蛋白水解级联的扰乱,及其下游信号传递作用,是肿瘤出现或肿瘤生长阻滞的基础。
关于 KLKs:该蛋白酶亚类的第一个成员是 Kraut, Frey 和 Werle 在 20 世纪 30 年代发现的,是一种蛋白水解酶,在希腊语中大量表达于胰腺激肽释放酶,因此被命名为组织激肽释放酶 (KLK1)。40 年后,在精液中寻找男性特异性抗原的研究在法医学上应用,这促使了前列腺特异性抗原的发现 (PSA; 后来改名为 KLK3)。迄今为止,KLK3 是特征最明显的激肽酶相关肽酶,也是临床实践中前列腺癌诊断和高危人群监测中最有价值的生物标志物。1989 年,对组织激肽释放素 (KLK1) 和 PSA (KLK3)的编码基因,以及对人类腺激肽释放素 - 1 (hGK-1; 后来改名为 KLK2)一个新的基因在人类染色体的 19q13.4 被映射到。
20 世纪 90 年代末,人们发现了 12 个编码激肽酶相关肽酶的新基因,根据它们的定位在 19q13.4,以及与前 3 个激肽酶基因的序列和结构相似性,人类基因组计划的测序数据显示,这些已经发现的基因与三种经典激肽释放酶定位在同一位点,即正常上皮细胞特异性 1 (NES1/KLK10),酶 / 蛋白酶 M / 神经蛋白 (PRSS9;KLK6),neuropsin / 肿瘤相关差异表达基因 - 14 (TADG14/KLK8), hippostasin / 胰蛋白酶样丝氨酸蛋白酶(TLSP / 人角质层胰蛋白酶(SCTE) KLK11)/KLK5) 和糜蛋白酶(HSCCE / PRSS6/KlK7)。对整个遗传位点的研究最终导致了另外 7 个以前未知的丝氨酸蛋白酶的克隆,将人类组织的 KLK 基因增加到 15 个,加上另外一个假基因 KLKP1。
这些基因在不同范围的组织中表现出不同水平的表达,它们表现出相当不同的表达模式。但根据最近建议的人类组织激肽释放酶命名法,每个基因都被称为了 KLK,后面跟着适当的数字,调控表达的蛋白质被称为 KLK,后面跟着适当的数字。
文献介绍:

了解完激肽释放酶相关肽酶 (KLKs),我们引入今天要介绍的这篇关于激肽酶相关肽酶对肿瘤微环境影响的综述,2022 年四月份发表在《nature reviews cancer》(if=60.716)上,详细介绍了 KLKs 对肿瘤微环境中的各种组分的影响机制。
一、肿瘤-基质的相互作用
肿瘤代表动态的组织微环境,除癌细胞和非细胞成分外,还包括各种细胞群 (基质细胞、内皮细胞、正常上皮细胞和免疫细胞)。尽管在肿瘤微环境中发现的所有细胞类型都不像上皮细胞和癌细胞那样强烈地表达 KLKs,但 KLKs 可以在免疫细胞中表达.
1.KLKs促进细胞外基质重塑(ECM)和细胞粘附信号传导
ECM 的改变是不同重构机制的结果,可分为四个主要过程:
(1) ECM 沉积,改变 ECM 组分的丰度和组成,从而影响 ECM 的生化和机械性能;
(2) 翻译后水平的化学修饰,改变了 ECM 的生化特性和结构特征 (图 1a);
(3) 蛋白降解,释放具有生物活性的 ECM 片段和 ECM 结合因子,可能需要释放细胞约束,如迁移障碍 (图 1b);
(4) 强制粘合的物理重构,通过对齐 ECM 纤维和打开细胞迁移通道影响 ECM 组织 (图 1c)。
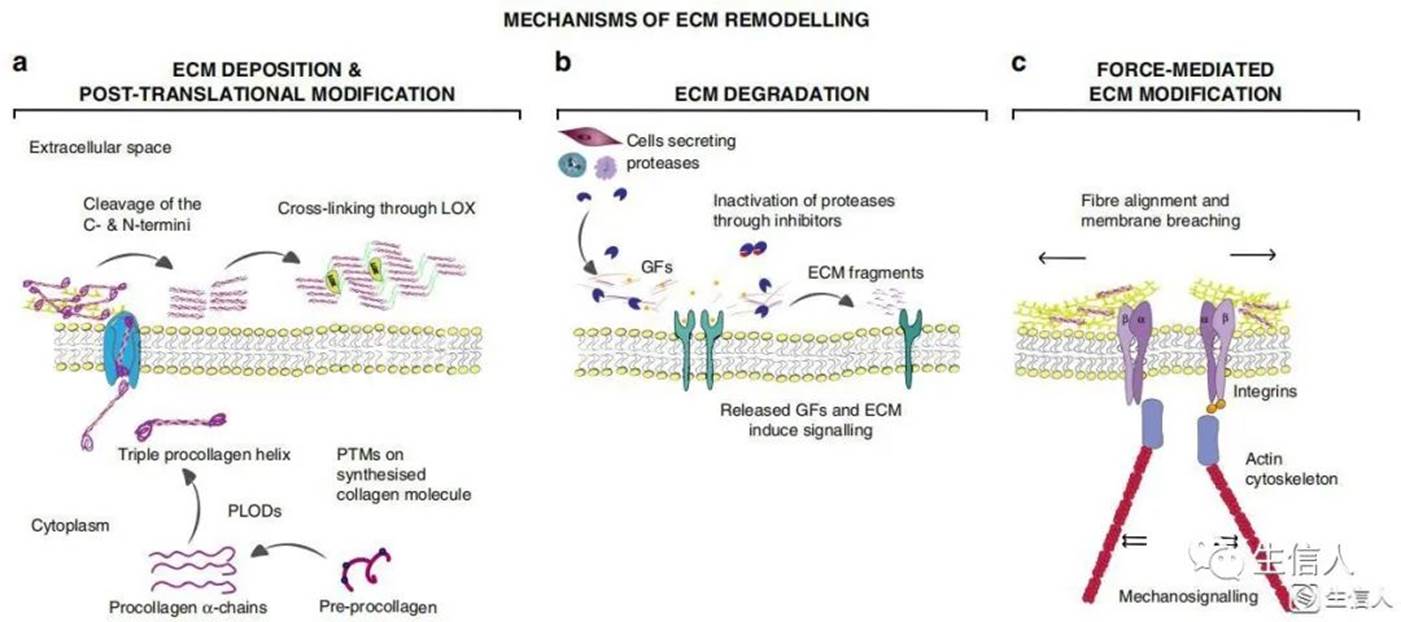
细胞外基质重塑 (ECM) 机制:
a ECM 沉积和修饰: 以胶原蛋白为例,前胶原蛋白被合成并转移到高尔基体,在那里它成为前胶原α- 链。这种前胶原分子经过多次翻译后修饰 (PTMs) 来改变其性质。PTMs 包括糖基化、前肽对齐、二硫键形成和羟基化。PLODs 对前胶原链的赖氨酸羟化使细胞内自发形成三螺旋并分泌到细胞外空间。在这里,c 端和 n 端上的前肽被蛋白酶切割,形成胶原原纤维。为了进一步组装胶原纤维,胶原原纤维由赖氨酰氧化酶 LOX 交联。
b ECM 降解: 包括基质金属蛋白酶在内的蛋白酶裂解 ECM 蛋白,释放基质结合生长因子 (GFs) 和细胞因子,以及包括基质因子在内的 ECM 片段,并清除细胞迁移的障碍。
c 力介导 ECM 的重构: 结合到 ECM 分子的整合素对 ECM 分子施加力。这可以改变 ECM 分子的构象,从而暴露结合位点来支持纤维的自组装,从而诱导纤维对齐。在这个修饰过程中,整合素施加的机械力也会导致基底膜的非蛋白水解性破坏,从而允许癌细胞侵袭。
KLKs 调节癌细胞与肿瘤微环境(TME)的相互作用

a
步骤 1:癌细胞分泌 KLKs
步骤 2:通过处理细胞间连接蛋白 (desmocollin (Dsc)/desmoglein (Dsg)) 控制癌细胞的凝聚。
步骤 3:粘附蛋白 (整合素、钙粘蛋白等) 的 KLKs 通过切割和调节癌细胞的附着和迁移。
步骤 4:klk 介导的细胞外基质 (ECM) 重构同时也影响了细胞与 ECM 的相互作用。
b
KLK 通过降解 ECM 蛋白底物包括纤维连接蛋白、层粘蛋白、胶原和富半胱氨酸蛋白 61(CCN)蛋白,改变 ECM 的组成并释放与 ECM 结合的生长因子。包括结合性表皮生长因子 (bEGF)、结合性成纤维细胞生长因子(bFGF)、血小板源性生长因子(PDGF) 和结合性血管内皮生长因子 (bVEGF) 成为可溶性(sEGF、sVEGF 和 sFGF),也释放可溶性 ECM 来源的生物活性片段。
KLKs 还参与许多隔离到 ECM 中的因子的激活,如肝细胞生长因子 (HGF) 和转化生长因子 -β (TGFβ)以及其他蛋白酶,如基质金属蛋白酶 (MMPs)、尿激酶纤溶酶原激活剂(uPA) 和 meprins。KLKs 介导的 ECM 重构对所有 TME 细胞群的影响。
2.KLKs在介导成纤维细胞(CAFs)与肿瘤细胞相互作用中的作用
KLKs 可能通过介导 CAF 和肿瘤细胞之间的串扰参与 TME 的重编程。在肿瘤发生的初始阶段,通常抑制肿瘤细胞生长的非 CAFs 被重新编程为支持肿瘤细胞生长的 CAFs。几名 KLK 成员已被确认可能参与了这次事件。例如,KLK4 在增生的前列腺上皮内瘤变病变中表达升高,这通常与前列腺癌的前体病变有关,也被认为是前列腺癌的前体病变,因此,KLK4 可能在早期肿瘤发展中促进邻近间质之间的相互作用。
除了调节肿瘤细胞和 CAFs 之间的相互作用,KLKs 通过作用于各种细胞间介质,包括生长因子,如细胞因子如白细胞介素,细胞外蛋白和 ECM 重构酶,促进活性基质的维持。

蛋白酶激活受体 (PARs) 的调控是 KLKs 调控的分子机制之一,参与了基质肿瘤细胞与上皮肿瘤细胞之间的串扰。PARs 是一个独特的 g 蛋白偶联细胞表面受体家族(包括 PAR1, PAR2, PAR3 和 PAR4),定位于癌细胞和肿瘤周围基质细胞。KLKs 可以在不同的位点水解 PAR1, PAR2 和 PAR4,激活不同的信号通路,称为经典或偏导信号通路,或相反,解除它们,使它们不可激活,所有这些都导致独特的细胞反应。
在 KLKs 水解蛋白之前, PARs 暴露了一个拴在结合自己的胞外受体的链接配体,并触发分子途径,如 MAPK 信号转导或 EGFR 的配体依赖性反式激活,调节生长因子的生物利用度, 促进癌细胞增殖。
在另一项研究中,用 KLK4 治疗永活的前列腺肌成纤维细胞 (WPMY-1) 可以诱导 PAR1 的激活和白细胞介素 - 6 (IL-6)的表达,这反过来刺激前列腺癌细胞的增殖和 KLK3、KLK4 的表达。最近,一项后续研究表明,KLK4 处理的 WPMY1 细胞在体外表达了 CAF 样表型标记物,如α-SMA、ESR1 和 SFRP1,以及一些被称为前列腺癌微环境调控蛋白的基因的高表达,包括赖氨酸氧化酶 (LOX),转胶质(TAGLN) 和成纤维细胞生长因子 1 (FGF1)。
其他 KLKs 调节 PARs,包括 KLK14,它对 PAR1、PAR2 和 PAR4 进行裂解,表明其他 KLKs 对基质细胞也有类似的作用。
除了调节肿瘤细胞和 CAFs 之间的相互作用,KLKs 通过作用于各种细胞间介质,包括生长因子,如上所述,细胞因子如白细胞介素,细胞外蛋白和 ECM 重构酶,促进活性基质的维持。KLK12 和 KLK14 可以通过紧密结合的潜伏期相关前肽 (LAP) 和潜在的 TGFβ结合蛋白 (LTBP) 复合物调节活性转化生长因子 -β(TGFβ)的生物利用度和释放(如上图),卵巢癌细胞中 KLK4、 KLK7 过表达可诱导 TGFβ1 信号转导。肿瘤细胞分泌的活性 TGFβ导致 CAFs 中 TGFβ信号通路的过度激活,从而使 TGFβ的功能可能随肿瘤环境而变化。例如,在上皮细胞中,在肿瘤起始过程中,TGFβ通过典型的 SMAD2/3 信号通路发挥肿瘤抑制因子的作用。同时,TGFβ也可以促进肌成纤维细胞分化,或通过激活 JAK/STAT3 通路诱导骨髓源间充质干细胞向 CAFs 分化。以及刺激癌细胞迁移和侵袭,表明 TGFβ在多种癌症特征中发挥关键作用。
二、癌症和免疫细胞的相互作用
显而易见,KLKs 是先天和获得性免疫的调节因子,具有促肿瘤或抗肿瘤的作用。KLKs 在诱导或抑制免疫应答方面的作用可能取决于对不同细胞类型中特定的细胞表面受体或细胞内靶点的激活,本文讨论了 KLKs 如何调节细胞因子、趋化因子或抗原呈递到适应性免疫系统细胞的功能,以及这种免疫调节功能对 TME 的潜在影响。
1. KLKs是免疫系统的靶点
诱导免疫反应涉及细胞和先天和适应性免疫系统的组成部分的合作努力。KLKs 通过诱导细胞毒性 T 淋巴细胞应答来调节抗肿瘤免疫应答,如 KLK4 和 KLK3,这是诱导抗肿瘤反应的重要过程。研究显示 CD8+ T 细胞能够破坏表达 KLK3 抗原的肿瘤,导致抗肿瘤反应。值得注意的是,小鼠缺乏内源性 KLK3 表达,因此,这种显著的细胞毒性免疫反应可能与其外源抗原性质有关。对卵巢癌抗原的免疫肽分析显示,KLK10 是人类白细胞抗原 I 类系统 86 的高递呈抗原之一,这表明 KLKs 是免疫治疗的潜在靶点。
大多数 KLKs 可以从其蛋白伴侣 (IGFBPs) 中释放可溶性生长因子,如胰岛素样生长因子(IGFs)。例如,KLK3 可以水解基质来源的 IGF1、IGFBP3 复合体,通过增加 IGF1 水平并调节良性前列腺增生衍生的纤维肌间质细胞生长。此外,KLKs 还能激活肝细胞生长因子激活因子(HGFA),激活 HGF 和诱发的体外浸润性癌细胞胰腺癌细胞株,通过 HGF 遇到信号级联。HGF 还可以增强胃癌细胞中 uPA 和 uPAR 的分泌,导致 ECM 重塑和细胞骨架的破坏,导致组织刚度降低,促进肿瘤细胞的迁移和侵袭。因此,KLKs 可能介导对组织刚度的相反作用,这取决于 TME 中的分子底物和相互作用的伙伴。深入研究 KLKs 在肿瘤间质相互作用中的作用,将有助于我们去更好地理解其在 TME 中的复杂背景作用。
2.KLKs作为免疫反应的调节因子

KLKs 通过细胞因子对癌症的影响:
(1) KLKs 可能有助于炎症细胞因子的形式转化为其活性可溶性形式,可作用于多种免疫细胞。例如,KLK1 以其激肽原活性而闻名,它能介导生物活性肽裂解缓激肽的释放,结合到 g 蛋白偶联受体家族激肽 B1 和 B2 受体 (B1R,B2R)。激肽是血管张力和通透性的重要调节因子,但也在炎症调节中发挥重要作用,包括调节细胞因子的产生。
(2) 并且,激肽刺激可触发乳腺癌细胞增殖,随后显示增加细胞 KLK6 和 KLK11 的水平,并降低 KLK10 的表达,导致侵袭性表型,这表明激肽也可以调控 KLK 的表达。cc 趋化因子配体 (CCL) 家族的几个成员的分裂也是由 KLK 直接蛋白水解或间接激活其他蛋白酶 (如 MMPs) 介导的,使它们成为具有炎症抑制作用的同源受体拮抗剂。
(3) 例如,CCL14 是具有 CCR1 或 ccr5 的白细胞的趋化剂和激活剂。KLK5、KLK8 可以打通八种氨基酸来解放 CCL14 的生物活性。KLK4、KLK7 表达的卵巢癌细胞被发现高浓度的 CXC-chemokine 配体 1。
(4) KLK3 介导一个抗肿瘤反应的细胞毒性 T 淋巴细胞活性的激活,同时它也可以通过补体 C5 蛋白的降解切割补体因子 iC3b (C3b s 因子 I 介导的降解片段)。用 iC3b 调理凋亡的肿瘤细胞,促进免疫细胞的清除,导致抗肿瘤免疫应答。因此,KLK3 对 iC3b 的裂解可能导致肿瘤效应。
(5) 此外,KLK14 可以从补体成分 C3 中产生 C3a 葡萄毒素,并诱导 C3a、c3ar 依赖性细胞信号通路,在小鼠足部水肿模型中发挥促炎作用。
(6) KLK4 则可以激活 PAR1,增加体外前列腺肌成纤维细胞 IL-6 蛋白表达,提示 KLK4 可以进行旁分泌调节,IL-6 是一种多效细胞因子,可维持体外前列腺癌细胞的促生存,并增加邻近癌细胞的 KLK 表达。KLK-PAR 信号也可能通过 NF-kB 信号通路促进肿瘤生长,导致炎症标志物表达增加,或阻止多聚 (adp - 核糖) 聚合酶 (PARP) 的裂解,并延迟支持免疫 T 细胞和 B 细胞存活的凋亡级联反应。因此,KLKs 可以通过外部互补依赖的蛋白水解机制,以及 PAR 信号传导,调节先天免疫反应的激活。这可能有助于肿瘤的致瘤性和转移。
(7) 由于一些经 KLK 处理的 ECM 成分具有趋化特性,因此 KLK 降解 ECM 可能对免疫系统产生二次影响。在角质形成细胞中,KLK5 和 KLK7 可以切割人类阳离子抗菌蛋白 (hCAP18) 的失活前体,这是在人类中发现的唯一一种抗菌肽,由多种免疫细胞产生。释放具有生物活性的 LL-37 抗菌肽。尽管没有证据表明这种活性在癌细胞中,LL-37 通过激活树突状细胞分化对其他免疫细胞具有趋化作用,最终具有促肿瘤的功能
(8) KLKs 也可能支持免疫细胞暴露在 TME 的恶劣环境下的生存,如缺氧、酸性条件和低营养水平。酸性 pH 已被证明可以通过抑制激肽释放淋巴上皮卡泽尔型相关抑制剂 (LEKT1),促进 KLK5 的高活性和下游的 KLK5-PAR2 级联,增强了内切顿综合征角化细胞的体外炎症过程。这种蛋白酶也被证明在免疫缺陷的体内乳腺癌模型中具有肿瘤抑制作用,通过抑制胆固醇合成的甲戊酸。
(9) 此外,酸性 ph 诱导的 KLK7 和 KLK8 表达可通过催化前列腺素生物合成和刺激炎症反应的环氧化酶 (COX) 途径增强体外胃癌细胞的侵袭能力。并且,在体外和体内的远处转移模型中,敲除 KLK5 或 KLK7 降低了三阴性乳腺癌细胞的转移并恢复了对 COX2 抑制剂的敏感性,表明这两个 KLKs 各自具有促肿瘤作用。
(10) 最近使用免疫标记或分子分析的研究表明,几种 KLKs (KLK1、KLK2、KLK4 KLK6 和 KLK8 KLK15)可以在体外循环分化的人类中性粒细胞和 HL-60 人类早幼粒细胞白血病细胞系 (主要包括前体细胞中性粒细胞) 中表达。中性粒细胞表达 IL-1β,其酶原形式可被 KLK7 激活。活性 IL-1β参与肿瘤介导的免疫抑制并促进转移,但这种功能与 KLK7 的联系尚未报道。
(11) KLK2、KLK5、KLK6 和 KLK9 在受促炎细胞因子刺激的中性粒细胞中表达,这表明 KLKs 在介导免疫应答中可能发挥作用。KLK1 可以在中性粒细胞上切割激肽片段,并在炎症部位产生中性粒细胞吸引肽,表明该分子在 TME 炎症部位的潜在作用。
(12) 在前列腺癌患者淋巴结转移组织样本的 T 细胞中发现了 KLK3 的表达,可能是通过肿瘤来源的癌细胞与 T 细胞之间的细胞外囊泡转移,提示 KLK3 可能通过调节 T 细胞功能发挥免疫调节作用。尽管目前对 KLKs 在肿瘤免疫环境中的作用了解有限,但这些蛋白酶在其他免疫介导的病理条件中已经被广泛表征。这表明 KLKs 可能通过多种方式参与免疫细胞功能归因于组织特异性底物或互补的蛋白水解调节机制。
三、血管生成和传播
KLKs 被认为通过控制生长因子的生物利用度、ECM 蛋白生成促进了血管生成抗血管生成肽或其蛋白水解活性的机制,参与肿瘤血管生成的调控。
1. KLKs调节促血管生成和抗血管生成因子的生物利用度
血管生成:

血管新生是由已有的血管形成新血管的过程。这是一个高度调控的过程,涉及内皮细胞 (ECs) 的迁移、生长和分化。这一调控机制在胚胎发育、伤口愈合和生殖中至关重要。然而,其任何调控途径的改变都可能导致代谢性疾病、心血管疾病、糖尿病视网膜病变、银屑病、系统性红斑狼疮、重要的是肿瘤生长和转移。
在无血管期,由于控制血管稳态的促血管生成因子和抗血管生成因子之间的平衡,肿瘤生长通常在大小上受到限制。并增加自身的血液供应,以提供足够的氧气和营养物质。这个过程被称为血管生成切换,从无血管状态进入血管生成阶段,对肿瘤生长和继续无限制增殖至关重要。因此,不像正常生理过程倾向于血管生成的负调控,肿瘤倾向于血管生成的上调。多种非互斥机制是肿瘤新生血管的主要参与者。这些包括芽生血管生成、非芽生血管生成、血管生成、血管生成模拟和肠套叠。然而,发芽的血管生成仍然是肿瘤细胞产生其血管系统的最被研究的机制。由于后者在肿瘤细胞生长、侵袭和外渗过程中的重要性,不同的血管生成抑制剂 (AIs) 已经被开发出来。

(1) 癌细胞分泌的 KLKs 作用于周围基质,调节旁分泌信号的传递,诱导促血管生成环境,触发活跃生长的血管网络的发展。这种血管生成开关涉及缺氧 TME 和缺氧诱导因子 1α (HIF1α) 途径促进的相关因素的复杂相互作用。HIF1α可以激活肿瘤细胞适应缺氧和血管生成相关基因的转录。非小细胞肺癌细胞暴露在缺氧环境下可诱导 KLK12 的表达,KLK12 在其 5 个侧翼区域中含有两个 HIF1α结合位点,表明缺氧和 HIF1α是 KLK12 表达的调控因子。
(2) 在心肌梗死模型中,KLK1 通过激活 B2 受体 (B2R) 诱导内皮细胞管形成,这导致了独立于血管内皮生长因子 A (VEGF-A)的 Akt / 内皮型一氧化氮合酶通路的激活。
(3) 另一个与诱导血管生成直接相关的 KLK 家族基因是 KLK12。首先,抗体抑制内皮细胞内源性 KLK12 活性,通过抑制激肽 B2R 信号通路,减少了内皮细胞的侵袭和毛细血管形态的形成,表明 KLK12 在激肽的产生中发挥了作用,但另一份报告显示,KLK12 并不直接生成激肽。
(4) KLKs 也可以通过裂解 lysa - 纤溶酶原, 产生抗血管生成的血管抑制素样片段,这在体外重组蛋白分析或内皮细胞系模型中观察到。从而证明了这些蛋白酶在 TME 中调节血管生成的功能复杂性。
(5) KLK3 抑制剂、KLK3 刺激肽和重组 KLK3 蛋白变体 (具有不同的蛋白水解活性) 的研究,使用人类脐静脉内皮细胞为基础的试管形成或发芽实验和在癌症背景下的体内研究,已经显示了 KLK3 的抗血管生成作用。
(6) KLK3 还可以阻断 VEGF 和 / 或 FGF2 介导的人脐静脉内皮细胞的增殖和迁移,使 KLK3 激动剂用于前列腺癌治疗合理化。
(7) 非活性 KLKs 的蛋白酶活性通常是在体外使用合成底物而不是与体内环境相关的全长底物进行测试。
(8) 宫颈癌细胞的代谢物可诱导内皮细胞分泌 KLK1,并伴随内皮细胞增殖和迁移的增加,但在内皮细胞条件培养液的刺激下则不能。这表明,内皮细胞释放的 KLK1 影响癌细胞诱导有丝分裂,促进血管生成。
(9) KLK4 和 KLK7 也可以切割血管稳定分子血栓反应蛋白 1 (THBS1)。
2. KLKs通过重塑血管而促进血管生成细胞外基质
(1) KLKs 调节许多促血管生成生长因子的生物利用度,如 VEGF121, TGFβ, FGF1,血小板源性生长因子和 MCP1 通过切割 ECM 和基质细胞蛋白,如纤维蛋白、纤维连接蛋白和富含半胱氨酸的蛋白 (CCN1 CCN6) 在体外癌细胞系和前列腺肌成纤维细胞模型中的表达,促进血管生成,骨骼和骨髓稳态和肿瘤发生。
(2) KLKs 还参与 uPA uPAR 系统的激活,这可能导致 ECM 重构,激活和 / 或释放促血管生成的,激活 MMPs12 的酶原形式和 KLK-PAR 。
(3) KLKs 也可以通过从纤维连接蛋白, 中产生促血管生成肽片段来诱导血管生成,损害可溶性纤维连接蛋白原纤维的组装; 因此,疏松的 ECM 促进了内皮细胞在体外的迁移。
(4) KLK12 在人肺内皮细胞形成的毛细血管周围的定位和大多数 KLK 能够劈裂纤维连接蛋白支持 KLK 在血管生成中的作用。因此,KLKs 可以影响 ECM 刚度,从而调节内皮细胞 VEGF-A 和 VEGFR2 表达的空间组织,并影响血管生成。
四、转移和治疗耐药性
如上所述,KLK 肽酶在破坏细胞 - 细胞连接、ECM 重构和激活前侵袭因子等方面发挥作用。这里,我们只强调它的功能影响 KLK 作用于调节恶性细胞的侵袭,导致转移性扩散并支持肿瘤在转移过程中的生长。KLKs 也在某些癌症的治疗耐药性中发挥作用。
1.KLK在肿瘤扩散和转移性扩散中的作用
在转移级联过程中,细胞 - 细胞和细胞 - 基质粘附的变化发生在转移的三个主要步骤: 癌细胞从原发肿瘤迁移——侵入周围组织——从血液或淋巴系统内渗和外渗,允许癌细胞循环到身体的所有器官。众所周知,KLKs 对 ECM 物理屏障的降解有利于这些过程,需要一个复杂的平衡,因为 ECM 的广泛降解干扰癌细胞的迁移和侵袭。
(1) 在肺癌细胞中转染 KLK13 和过表达 KLK13 增加了小鼠肺癌体内异种移植模型的肺转移,可能是通过层粘连蛋白的切割和 N - 钙粘蛋白表达的诱导,在体外模型中可以看到,KLK13 被诱导过表达,而沉默 KLK13 逆转了这些。与正常肺细胞系相比,肺癌细胞中 KLK13 表达的增加归因于其上游启动子的去甲基化,这在体外用去甲基化剂处理正常肺上皮细胞时得到证实。提示这可能是 KLK13 诱导肺癌的机制。相反,KLK13 的转染和过表达也与口腔鳞状细胞癌细胞系中肿瘤细胞运动能力的降低以及粘附分子桥粒胶蛋白 2 (Dsc2)、桥粒蛋白和 E - 钙粘蛋白的高水平有关。
(2) 同样,KLK6 在高 KLK6 表达的胃癌细胞系克隆与低 KLK6 表达的胃癌细胞系克隆相比,KLK6 在体外具有不同的作用,可能是通过在胃癌患者组织中观察到的较高的 MMP 和 VEGF 表达介导的。KLK6 转染和过表达也通过 TGFβ和 SMAD 信号通路增加了结肠癌细胞的体外侵袭能力。通过转染 KLK6 水平升高的乳腺癌细胞系发现,在免疫缺陷的乳腺癌小鼠模型中,KLK6 过表达增加了原发肿瘤的生长和转移。然而,生理水平的表达表现出相反的作用,通过抑制肺转移,提示浓度依赖的作用。
(3) 因此,KLKs 在肿瘤中可能具有多方面的作用,当在生理水平上表达时,但当过度表达时,如在特定癌症患者中观察到的那样,KLKs 可能会促进肿瘤进展,反之亦然。
(4) KLK 介导的 PAR 依赖性在结肠癌、肺癌、口腔鳞癌和前列腺癌细胞中诱导癌细胞迁移。此外,KLK4 和 PAR2 在前列腺癌骨转移灶的共同定位结果提示 KLK 介导的 PAR 依赖性作用也可能发生在转移灶。KLK2 和 KLK4 也可以诱导纤溶酶原 uPA、uPAR 轴,通过纤溶酶连接这三个蛋白水解系统导致 MMP 活化。事实上,uPA 是 KLK4 的第一个识别的底物,并且在 KLK4 编码质粒转染的卵巢癌球形模型中上调。因此,探索 KLKs 在转移过程中的介导作用可能进一步使靶向这些蛋白酶在癌症治疗中的作用成为可能。
2.肿瘤转移扩散中的KLKs
骨转移在许多癌症中都很常见。在这方面,KLKs 激活 TGFβ信号的能力可能在原发肿瘤部位之外产生影响,并在调节骨稳态方面产生影响。在前列腺癌和成骨成骨样细胞共培养模型中,TGFβ1 介导 KLK4 上调。这提示了在前列腺癌骨转移中观察到的高 KLK4 水平可能的机制。TGFβ也被称为免疫抑制的介质,并通过多种机制促进肿瘤细胞迁移和骨转移。因此,KLK-TGFβ的激活可能在这些转移位点创建一个免疫抑制利基,促进肿瘤进展并可能通过影响肿瘤细胞功能促进肿瘤细胞侵袭和转移。
癌细胞表达的整合素可与成骨细胞表达的骨桥蛋白 (OPN) 和玻连蛋白结合,并定植到靶组织。整合素在卵巢癌细胞中的表达受 KLK4、 KLK7 的表达调控,最近,OPN 被认为是 KLK7 在卵巢癌细胞中的新底。癌细胞分泌的 KLKs 也间接调节促进骨定植的细胞因子受体的表达。细胞因子受体,由 RANKL 和 MCSF1 控制的破骨细胞介导的再吸收导致骨材料溶解和有机基质暴露于蛋白酶,包括 KLKs、MMPs 和组织蛋白酶,释放生长因子,导致局部成骨形成。生长因子的 IGF 家族的生物利用度,可能在前列腺癌骨转移中起着有争议的作用,受几乎所有 KLKs 的调控。
3.KLK在治疗耐药性中的作用
一些研究已经证明了 KLK 表达与治疗反应之间的相关性,并确定了不同的机制。体外实验中,卵巢癌细胞中 KLK4 和 KLK7 单独表达的增加通过促进多细胞聚集的形成促进了化疗耐药,这是一种已知发生在卵巢癌腹水中的适应性反应,可能分别通过α5/β1 整合素途径和 uPA 机制进行。同样,KLK6 通过激活 PAR1 和 PAR2,在体外促进胶质母细胞瘤细胞对放疗诱导的凋亡的抵抗。在体外和小鼠胃癌模型中,KLK6 的表达也降低了 aurano 芬耐药诱导的细胞死亡,增加了自噬介导的 aurano 芬耐药,提示 KLK6 通过自噬调节可能增加化疗耐药。
据先前的研究发现,KLK4 可调节 AR 和 mTOR 信号通路来调节前列腺细胞的生存,在体外实验中,KLK4 的下调可使前列腺癌细胞对凋亡诱导药物敏感,提示沉默 KLK4 可能有显著的治疗效果。KLKs 也可以通过其他机制促进化疗耐药,例如通过诱导上皮间充质转化 (EMT) 过程,减少肿瘤细胞粘附,增加 TME 中的侵袭性,或在低氧条件下增加组织刚度,这可能影响药物外膜率并促进化疗耐药。例如,在头颈部鳞状细胞癌细胞中沉默 KLK6 的表达增强了 EMT 特征和β-catenin 的积累,导致体外肿瘤细胞的增殖和活力降低,并提高了对辐照的抵抗力 55。因此,KLK 功能的恢复可能有助于 EMT 信号通路的药理靶向。上述研究表明,KLKs 在介导治疗性耐药中具有复杂的细胞特异性作用,这些机制部分重叠,可以作为未来研究的一个关键领域,以支持可能的新治疗干预。
五、靶向KLK治疗方法

多项研究已经证明了 KLKs 在癌症进展中的临床意义和生物标志物作用,KLKs 的表达失调和参与癌症的各种特征,使它们成为癌症治疗的药理靶点。KLK 靶向药物可大致分为:
1. 抑制剂 (天然、蛋白、多肽和小分子):不可逆的紧密结合内源性抑制剂,如丝氨酸、kazal 型抑制剂、蛋白类抑制剂和体液中的其他调节 KLK 蛋白水解活性,并维持细胞稳态的抑制剂。这种调节机制的失效可能导致几种病理生理状况。在癌症背景下,在卵巢癌患者腹水中观察到丝氨酸蛋白酶抑制剂和 KLK 抑制剂复合物。这些天然抑制剂和内源性阳离子(如 Zn2+) 已被认为是前列腺癌的潜在治疗方法。此外,重新设计和工程内源性和天然的 KLK 抑制剂,如丝氨酸蛋白酶抑制剂,以增加其靶向 KLK 的特异性,已经引起了相当大的药理学兴趣,其中一些正在进行人体临床试验。
2. 可激活的前药物:天然抑制剂包括组织特异性 KLK 活性也被用于设计前药物,为了在临床试验中增加这些前药物的疗效,最近的研究通过改变 KLK3 肽底物序列或使用 ph 敏感的纳米复合物来探索这种前药物的修饰,从而导致更快更有效的释放和提高抑制剂中的活性,此外,自然毒素在表达 KLK 的肿瘤组织部位激活和杀伤活性。例如, 一种具有 KLK3 抗原特异性序列的原溶血素 (PRX302) 在体内小鼠和猴子模型中表现出显著的溶细胞活性,并在半脾肝转移模型中被探索用于靶向治疗转移性前列腺癌。
3. KLK 靶向免疫治疗:由于一些前列腺 KLKs 的表达受 AR 信号调节, KLKs 特异性抗体,包括 KLK2 和 KLK3,已经被开发用于准确测量肿瘤相关的 KLK 表达,反映 AR 驱动的表达变化,在前列腺癌患者使用 AR 轴靶向药物治疗后。最近,基于 KLK 的单克隆抗体已被用于放射治疗靶向和治疗目的。KLK3 作为肿瘤相关抗原诱导对表达 KLK3 的前列腺肿瘤细胞的限制性免疫反应和抑制肿瘤生长的能力被用来开发一种 PROSTVAC 疫苗。由于无法区分表达 KLK3 的正常组织和肿瘤组织,最近的临床试验并没有支持 PROSTVAC 的总生存期的改善。尽管在多个小鼠和非人灵长类动物模型中进行了临床前评估,但没有一种 KLK 靶向制剂成功转化为临床。这可能是由于 KLKs 的生物学和表达在人与动物模型之间存在差异,因此这些发现的翻译相关性需要进一步研究。利用人源杂交临床前小鼠模型可能为了解 KLK 靶向抑制剂的免疫耐受机制和规避免疫耐受的新策略提供额外的见解。另一种更近期的方法是利用长非编码 RNAs,小干扰 RNA 引起 RNA 干扰和通过 microRNAs 来调节 KLK 的表达。然而,这些尚未进入临床试验。
4. KLK 功能恢复:由于广谱 MMP 抑制剂在临床试验中失败,由于抑制剂的选择性差和不同 MMP 的抗肿瘤作用没有得到很好的研究,开发蛋白酶靶向治疗的总体需要谨慎执行。因此,许多抑制剂同时抑制促肿瘤蛋白酶和抑制癌症发展或具有重要生理功能的蛋白酶,这也是考虑 KLK 抑制剂治疗时应考虑的因素。
参考文献:
Srinivasan S, Kryza T, Batra J, et al. Remodelling of the tumour microenvironment by the kallikrein-related peptidases[J]. Nature Reviews Cancer, 2022: 1-16.
小编说:
在许多癌症的病理生理学中,KLKs 是关键的效应因子。KLKs 可能具有促肿瘤和抗肿瘤功能,了解其在各种相互作用的蛋白水解和信号级联反应中的作用,以及底物的生物利用度和定位,这些决定了其在癌症发展、进展和转移中的作用。这篇综述概述了 KLKs 的自分泌和旁分泌作用如何根据 TME 中的刺激或信号通路的失调而变化。一些 KLKs 被用作生物标志物,KLKs 在化疗和放疗耐药中的作用和在免疫治疗中的潜力也提示了其在肿瘤治疗中的关键作用。从转化和有效靶向治疗的角度来看,KLK 研究领域将应用这些更复杂的分析,以提高对 KLK 在癌症进化和进展中的功能重要性和潜在分子机制的理解,以及它们的治疗潜力。
转载自公众号《生信人》,如有侵权,请联系删除。

